인체 내의 활성산소는 슈퍼옥사이드라디칼에서 시작하여, 과산화물로, 그리고 다시 과산화수소로 그 해로운 연결관계를 이어간다. 조금씩 강도를 약화시켜 가는 과정이라고 볼 수 도 있겠다. 가장 치명적인 수산화라디칼을 직접적으로 제어할 수 있는 방법을 인체는 가지고 있지 않다. 다만 그 전구체이자 매개체인 과산화수소를 무해한 물로 전환시켜 미리 사전에 대처하는 방법만이 존재할 뿐이다. 이제 활성산소의 체인을 따라 그 출발점부터 인체가 이들을 어떻게 처리하는지 그 관련된 효소들의 활약을 순차적으로 짚어보자.
생리학적 조건에서 상대적으로 안정적이고 생물학적으로 중요한 다양한 화합물들과 반응하는 과산화수소는 다양한 생체분자와 상대적으로 느리게 반응하기 때문에 독성이 약한 것으로 알려져 있다. 활성 라디칼들의 제거 과정은 주로 과산화수소가 중심이 되는데 쉽게 분해가 잘되는 그 본성적 특징을 활용하기 위함이 아닐까 짐작한다. 활성산소의 출발점인 슈퍼옥사이드라이칼 역시 일단 과산화수소로 전환시킨 후 다음 단계로 넘어간다.
1단계 슈퍼옥사이드라디칼을 과산화수소로 변환.
슈퍼옥사이드불균등화 효소(SOD)
슈퍼옥사이드라디칼이 과산화수소로 전환되는 과정을 촉진시켜 주는 효소이다. 이름이 모든 것을 설명한다. dismutase는 dismutation(불균등화)을 의미하는 동사에 효소를 의미하는 접미사 '-ase' '-ase'가 붙어있다. 불균등화란 하나의 물질이 산화와 환원반응을 동시에 일으켜 그 결과로 각각 다른 생성물을 만들어내는 것을 가리킨다. 자연 상태에서는 이러한 반응이 천천히 일어나지만 촉매를 이용하면 그 반응 속도가 훨씬 빨라지도록 만들 수 있다. 즉, 2개의 슈퍼옥사이드라디칼과 2개의 양성자가 동시에 산화와 환원 반응을 일으켜 과산화수소와 산소를 만들어 내는데, 이 반응을 빠르게 가속화하는 촉매가 Superoxide dismutase(이하 SOD)인 것이다.

SOD 패밀리
SOD 패밀리에는 SOD1, SOD2, SOD3라는 3개의 이소형(isoform)이 있다. 이소형이란 하나의 같은 유전자로부터 만들어졌으나 그 구조나 기능이 완전히 같지는 않은 단백질, 효소등을 의미한다. SOD1은 세포질에, SOD2는 미토콘드리아, 그리고 SOD3는 세포외부, 뇌척수액과 뇌혈관 등에서 발견된다. SOD는 거대하고 복잡한 단백질 구조를 가지며 직접적인 역할을 하는 그 활성부위에 금속이온을 갖는데 이 금속이온들이 전자의 이동을 도와 촉매활동을 한다. SOD1과 SOD3은 그 반응의 중심에 각각 구리와 아연을, SOD2는 망간을 포함한다.
금속이온의 역할(구리, 아연, 망간 등)
이 반응에서 금속이온의 역할[1]을 구체적으로 이해하기 위해 SOD1의 메커니즘을 간단하게 들여다보자. SOD1에는 구리이온이 관여한다.
먼저 1단계로, 슈퍼옥사이드라디칼 한 분자가 구리이온에게 전자를 주고(산화)하여 산소분자가 되고, Cu2+는 전자를 획득(환원)하여 Cu가 된다. 이때, Cu와 결합되는 활성 부분의 아미노산 잔기인 히스티딘과 Cu이온과의 결합이 끊어지면서 His63 Nε1(63번 위치에 있는 히스티딘의 측면사슬에서 첫 번째 위치의 질소를 의미)이 양성화된다. 양성화란 원자나 분자, 이온에 수소 양이온이 부착하는 것을 의미한다.
2단계로, His63 Nε1부터 받은 수소 양성자와 Cu로부터 전자 하나가 슈퍼옥사이드라디칼에 공여된다. Cu는 전자를 공여하고 산화되어 Cu2+로 돌아가고, 슈퍼옥사이드라디칼은 전자를 하나 획득하여 환원된다. 끊어졌던 히스티딘과의 bridging 결합은 원래대로 복원된다. 금속이온의 역할이 매우 중요하다는 것을 알 수 있다. 그 말은 우리가 라디칼을 체내에서 제대로 제거하기 위해 구리, 아연, 망간과 같은 미네랄을 적절히 섭취해야 함을 시사한다.
SOD가 슈퍼옥사이드 라디칼을 일단 과산화수소로 전환시켰다. 사실 썩 만족스러운 안전장치라 할 수없다. 그래서 다음단계로 과산화수소 처리에 필요한 효소들 중 대표적인 2가지를 살펴보자.
2단계 과산화수소를 물로 변환.
과산화수소는 이전글에서 살펴보았듯이, 슈퍼라디칼음이온으로부터 유래하여 생성된다.

또한 위에서 살펴본 것과 같이 슈퍼라디칼이 SOD에 의해 촉매 되어 과산화수소로 형성되기도 한다. 우리 인체는 라디칼을 일단 과산화수소로 만들어 놓고, 이를 다시 무해한 물로 전환시켜 라디칼의 영향을 무력화시키는 방법을 쓴다. 이 해독 작용에 투입되는 여러 항산화 효소가 있다. 하지만 이 중에서도 가장 강력하고 가장 빠르게 작용하는 것으로 알려진 효소가 바로 글루타티온 퍼록사이드(Glutathione Peroxidase), 즉 GPx이다.
1. 글루타티온 퍼록사이드(Glutathione Peroxidase, Gpx)
GPx 효소는 라디칼을 물로 빠르게 전환시키는 효소로서, 이 효소의 작용에는 셀레늄과 글루타티온을 반드시 필요하다. 글루타티온은 글루탐산(Glutamate), 시스테인(Cysteine), 글리신(Glycine)의 3가지 아미노산으로 구성된 우리가 이미 잘 알고 있는 강력한 항산화제이다. GPx는 우리 신체의 다양한 기관 즉, 간, 적혈구, 신장, 두뇌등의 조직 내 세포질, 미토콘드리아와 퍼옥시좀에서 발견된다.
GPx의 활성부위에는 셀레노시스테인(selenocysteine, Sec)이 포함되어 있다. 이 부분이 촉매작용의 핵심부위로서 매우 중요하다. 이 셀레노시스테인은 우리가 많이 접하는 시스테인 아미노산과 아주 흡사한데, 사실 황(S) 자리에 셀레늄(Se)이 대신 들어간 형태이다. 시스테인이 그 구조안에 S원자와 수소(H) 원자가 결합한 티올(S-H, thiol) 작용기를 갖듯이, 셀레노시스테인 역시 셀레늄(Se) 원자와 수소(H) 원자가 결합한 셀레놀(Se-H, Selenol) 작용기를 갖는다.
1. GPx효소의 셀레놀이 라디칼과 먼저 반응한다. 지질과산화물(ROOH) 또는 과산화수소(H₂O₂)로부터 산소 원자를 하나 받아 Se-H(셀레놀)가 Se-OH(셀렌산)이 된다. 이때, 산소 원자 하나를 잃은 ROOH는 알코올(ROH), H₂O₂는 H₂O로 해독된다.

2. GPx의 셀렌산(Se-OH)의 OH는 GSH로부터 받은 수소 한 원자와 결합하여 H₂O를 생성하고, 이들은 글루타티올화된 셀레놀(Se-SG)을 형성한다.
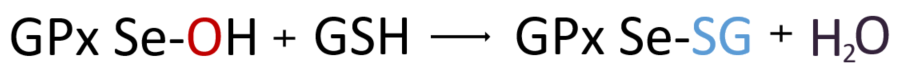
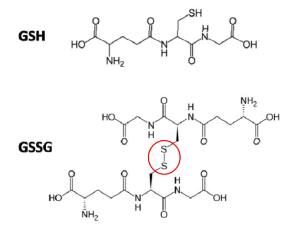
3. 여기에 다시 한분자의 GSH가 내어 놓은 수소를 받아 Se-H로 돌아감으로써 GPx는 다시 효소작용을 재개할 준비가 된다. 이때 차례대로 수소를 내어놓은 두 분자의 GSH 이황화결합을 하여 GSSG, 즉 산화된 글루타티온이 된다.

1-1 글루타티온 환원효소 Glutathione Reductase(GR)
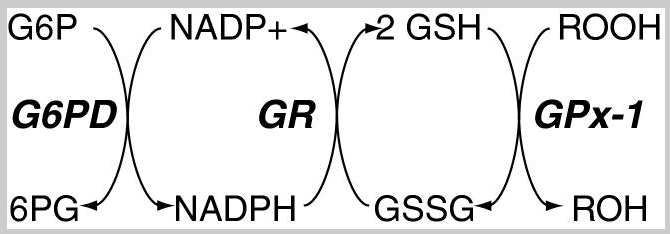
GSH가 라디칼을 중화하고 GPx효소를 원위치로 돌려놓고 본인은 산화하여 산화된 글루타티온인 GSSG이 되었다. 가장 강력한 항산화제인 GPx를 끊임없이 재가동하려면 우리는 산화된 GSSG를 다시 환원시켜 원상 복구하여 다시 항산화활동에 재투입해만 할 터인데 다행히도 인체는 이를 위해 Glutathione Reductase(GR)라는 글루타티온 환원효소를 보유하고 있다. GSSG를 환원시키기 위해, 즉 전자를 받을 곳이 필요한데 이를 위해 NADPH가 등장한다. 즉 GR촉매효소의 핵심은 전자공여체 NADPH이고, 이는 해당작용(glycolysis)에서 분기(shunt) 되어 해당작용과 평행하여 진행되는 대체 경로라 할 수 있는 HMP 션트(hexose monophosphate shunt)에서 만들어진다 [2]. 참고로, 오탄당 인산 경로(pentose phosphate pathway :PPP)라고도 불리는 이 경로를 이용해 인체는 글루타티온 항산화제의 계속적인 항산화활동을 가능케 해 줄 전자공여체 NDHPH의 생산과, DNA와 RNA 분자를 구성하는 뉴클레오티드 합성을 위한 리보스-5-인산을 생성한다. 6 탄소 설탕인 포도당(C6H12O6) 은 해당 시점에 세포의 개별적 요구에 따라 해당과정 경로로 들어가거나 대체 HMP 션트로 들어갈 수 있다.
글루타티온 환원효소(GR)는 FAD(플라빈 아데닌 디뉴클레오티드)가 반드시 있어야 작동하는 단백질이다. NADPH가 먼저 FAD에 전자를 주고, 환원된 FADH2는 GR의 활성부위에서 GSSG 내로 전자를 전달하고, 이것으로 인해 GSSG 내의 두 개의 시스테인 잔기에 수소 양성자가 환원되어 두 분자의 GSH가 생성되어 GSH로 복구된다. 이렇게 GR은 한 분자의 GSSG를 두 분자의 GSH로 환원시키는 촉매로서 세포 내 환원 및 산화된 GSH의 비율을 유지하는 중요한 역할을 한다.
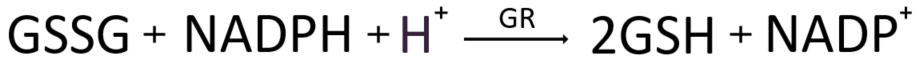
참고로 FAD는 필수비타민 B2로 잘 알려진 리보플라빈으로부터 생성되는 것이고, NAD와 NADPH는 비타민 B3 니아신으로부터 생성된다. 리보블라빈은 생체에서 합성되지 않으며, 니아신은 아미노산 트립토판으로부터 전환되어 만들어 낼수있지만 그 양이 충분히 보장되지 않으므로 음식이나 기타 다른 방법을 통해 충분히 보충할 필요가 있다.
GSH:GSSG 비율
글루타티온이 ROS를 해독하는 큰 그림과 각 단계별로 필요한 효소들의 모습이다. 우리 인체는 이러한 효소들을 끊임없이 항산화활동을 재생할 수 있도록 도와준다. 환원된 형태의 글루타티온인 GSH와 산화된 형태의 글루타티온 GSSG의 비율은 세포건강을 암시하는 지표가 될 수있으며 [4], 정상적인 세포 환경에서는 GSH가 훨씬 높은 농도로 존재한다. 즉, 정상적인 세포 환경에서는 GSH:GSSG의 비율은 100:1 정도이나 산화스트레스 상황에서는 이 비율이 10:1, 심지어 1:1까지도 떨어져[5] 이 비율을 통해 산화스트레스의 정도를 가늠할수 있는 척도가 되기도 한다. GSH가 낮게 나타난다는 것은, 산소라디칼의 증가, 항산화 능력의 감소, 또는 글루타티온 재활용 매커니즘상의 장애가 있음을 암시할수있다. 이 둘간의 비율을 모티텅링함으로써 세포의 산화스트레스로 인해 발생된 손상과 관련 질병에 대한 민감도를 평가하는데 유용한 정보를 제공할수있다.
2. 카탈라제(Catalase)
세포내 기관인 퍼옥시좀(peroxisome)에 가장 고농도로 존재하는 과산화수소 해독 효소이다. 퍼옥시좀에는 과산화수소를 생성시키는 반응들이 다양하게 일어나는 곳이며 대표적인 예로 지방산 분해를 들 수있다. 많이 발생하는 과산화수소의 균형을 유지하기 위해 퍼옥시좀에서 카탈라제가 많은 양으로 관찰되는 것은 합리적으로 보인다. 물론 카탈라제는 세포밖에서도 발견된다.
다양한 효소들이 과산화수소의 농도를 낮추기 위해 활동하지만 만약 그 농도가 1 mM(마이크로몰)이상이 될 시에는 카탈라제가 주요 방어효소로 맹활약하게된다[6]. 종종 산화 스트레스 상황에서 이 정도의 농도를 볼 수있다.
카탈라제는 500개가 넘는 아미노산이 연결된 폴리펩타이드가 4개인 거대한 4량체 단백질이다. 이 4량체는 각각 그 하부 단위로 과산화수소와 직접 반응하게 될 4개의 heme group을 포함하고 있다. Heme 그룹은 porphyrin 링 구조에, 그 중앙에 철(Fe3+)을 갖고 있어 이 부분이 조효소로 역할하여 산화 및 환원 반응을 통해 과산화수소 사이에서 전자의 이동이 생기고 이 과정을 통해 최종적으로 두 분자의 과산화수소를 두분자의 물분자와 하나의 산소분자로 분해한다. 아래 이미지에서 4개의 주황색 점이 카탈라제 촉매작용의 활성 부위인 heme group이다.

| ☞ Heme group은 헤모글로빈의 한 부분으로 잘 알려져 있다. 조직에 산소를 공급하는 일은 적혈구 내의 헤모글로빈(Hemoglobin)이 수행한다. 헤모글로빈은 글로빈 단백질에 헴(heme)그룹이 부착된 구조를 가지는 4량체 금속단백질이다. 하나의 산소분자가 하나의 heme group 중앙의 철에 결합할 수 있으므로 한번에 4개의 산소분자를 수송할수 있다. 참고로 근육에 산소를 공급하고 저장하는 마이오글로빈(Myoglobin)은 하위단위가 1개라는 점이 차이다. 여기에서 글로빈 폴리펩타이드 단백질에 부착된 heme group을 Prosthetic group이라고 부른다. Prosthetic group(보결분자단)이란, 아미노산으로 만들어진 단백질은 아니지만 다른 단백질에 영구적으로 단단히 부착되어 그 단백질의 효소 촉매 작용, 구조적 안정성등에 굉장히 필수적인 역할을 하는 것들을 말한다. 우리에게 잘 알려진 대표적인 Prosthetic group으로는 heme group, FAD, FMN, 코엔자임 A(CoA)등이 있다. |
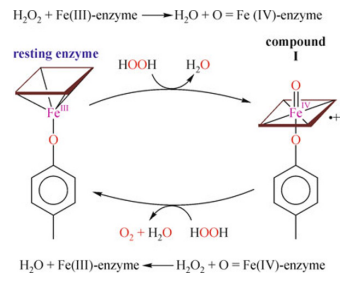
카탈라제 효소 반응의 구조
1단계: Cpd I(compound imtermediate I) 중간체 형성
첫번째 과산화수소(H2O2)가 효소와 결합하면, 효소 활성 부위의 heme group이 과산화수소분자에게 두 개의 전자(F3+와 포르피린 고리로부터 하나씩)를 제공한다. 그 결과로 산소페릴 π-양이온 포르피린 라디칼(oxoferryl π-cationic porphyrin radical) , 또는 Cpd I이라고 부르는 중간 화합물이 형성된다. 효소의 활성 부위 내 히스티딘 잔기를 통하여 과산화수소의 양성자(H+) 하나가 한쪽 끝에서 떨어져나와 반대쪽 끝으로 이동하게 되는데, 이 과정은 극성(polarization)을 만들어 내게 되어 과산화수소의 산소 O-O 결합을 깨뜨리게 되어 결과적으로 하나의 물 분자를 생성하게 된다.
2단계
두번째 과산화수소가 Cpd I에 두개의 전자를 내어주고 환원시켜 원래의 Fe3+ 상태로 복원시키고, 물 한분자와 산소 한 분자를 방출한다.
그래서 결과적으로 이를 화학적으로 표현하자면,
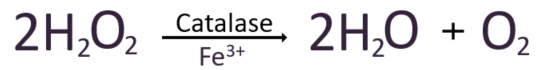
활성산소를 무해한 물로 분해하여 산화스트레스를 방지하는 역할을 하는 기전과 그 중심에 있는 대표적 항산화효소들을 살펴보았다.
[참고 자료]
[1] Perry JJ, Shin DS, Getzoff ED, Tainer JA. The structural biochemistry of the superoxide dismutases.
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3098211/
[2] Biochemistry, Hexose Monophosphate Pathway
https://www.ncbi.nlm.nih.gov/books/NBK551687/
[3] Glutathione Peroxidase-1 in Health and Disease: From Molecular Mechanisms to Therapeutic Opportunities
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3159114/
[4] Redox status expressed as GSH:GSSG ratio as a marker for oxidative stress in paediatric tumour patients
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3506742/
[5] Redox status expressed as GSH:GSSG ratio as a marker for oxidative stress in paediatric tumour patients
https://www.spandidos-publications.com/10.3892/ol.2012.931#b12-ol-04-06-1247
[6] Monofunctional Heme-Catalases
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC9687031/
Thirty years of heme catalases structural biology
Human catalase: looking for complete identity
https://link.springer.com/content/pdf/10.1007/s13238-010-0113-z.pdf
'활성산소(ROS)관련' 카테고리의 다른 글
| ROS: 활성산소에 대한 나의 정리 (0) | 2024.03.10 |
|---|---|
| ROS: 항산화물질: 항산화네트워크(Antioxidant Network) (0) | 2024.03.10 |
| ROS: 자유라디칼은 필요악(necessary evil)인가. (0) | 2024.03.10 |
| ROS: 신호전달물질로서의 ROS (1) | 2024.03.10 |
| ROS: 활성산소 생산이 목적인 호흡폭발(Respiratory burst) (0) | 2024.03.10 |